When we last left off, we were feeling a little bummed. Gregor Mendel never received the recognition he deserved for discovering the basic laws of heredity, at least not while he was alive. August Weismann was struggling against criticisms of his theory, which said that inheritance passed from parent to child solely through gametic cells in a process known as fertilization that was essentially meiosis in reverse. And it was clear that had Weismann (or anyone else, for that matter) just known about Mendel’s work, the burgeoning field of genetics would be poised to take a giant leap forward. Why’s that? Because Weismann and others had observed that, during fertilization, chromosomes come together from both parents in a way that was very similar to Mendel’s theoretical explanation of how heredity in pea plants worked. Mendel lacked the intimate knowledge of chromosomes that Weismann had, but Weismann lacked the knowledge of heredity that Mendel had. Once the two pieces of the puzzle were put together, it would be clear: the inheritance of genetic traits from one generation to the next is determined solely by the chromosomes that interact when a sperm and an egg unite.
The resurrection of Mendel
We rejoin the tale in the 1890’s in the company of three scientists. The first, Hugo de Vries, a botany professor at the University of Amsterdam, was fast approaching his 50th birthday and was already established in the field: he coined the terms ‘gene’ and ‘mutation,’ among other achievements. The second, Carl Correns, was a thirtysomething tutor at the University of Tubingen, still trying to crack into the ultracompetitive world of academic research. The third, Erich Tschermak von Seysenegg (I have no idea how to pronounce that), was not a geneticist, unlike the other two. He was a plant breeder working for an agricultural business, fresh out of school and with hopes of breeding hardier and more productive crops. All three men worked feverishly toward the answer to a deceptively simple question: Why is it that plant breeding experiments seem to produce offspring with traits that follow simple mathematical rules?
All three were converging on the same answer: Mendel’s answer. The details are disputed by historians of science, but at some point in the late 1890’s, they each became aware of the existence of Mendel’s work on pea plants. De Vries published his work first: in late March of 1900 in the journal of the very prestigious French Academy of Sciences, Comptes Rendus. However, nowhere in his paper did he mention Mendel’s work on pea plants some 35 years earlier. Correns published his paper a few months later in the somewhat less prestigious German Botanical Society’s journal. In it, he called de Vries out on his omission. From the English translation of Correns’s paper:
…the same thing happened to me which now seems to be happening to de Vries: I thought that I had found something new. But then I convinced myself that the Abbot Gregor Mendel in Brünn, had, during the sixties [ed: the 1860’s, that is], not only obtained the same result through extensive experiments with peas, which lasted for many years, as did de Vries and I, but had also given exactly the same explanation, as far as that was possible in 1866.
Tschermak, publishing in the same journal as Correns in June of 1900, seemed to feel the same way:
The simultaneous “discovery” of Mendel by Correns, de Vries, and myself appears to me especially gratifying. Even in the second year of experimentation, I too still believed that I had found something new.
Each of the three claimed to have come up with Mendel’s laws before they read Mendel’s work. Historical evidence suggests that Tschermak didn’t fully understand what he had discovered, and most discount young Tschermak as a true “rediscoverer.” (That being said, this opinion changes about once a decade, as historical opinions are wont to do.) The case of Correns is pretty clear-cut, too. Most historians agree that Correns did, in fact, rediscover the laws of genetic inheritance before becoming aware of Mendel’s paper. De Vries has always been the big question mark. There is significant historical evidence that de Vries attempted to cover up the influence that Mendel’s work had on his own thinking, while repeatedly asserting that his discoveries were independent of everyone else’s. One of these pieces of evidence was pointed out by Correns himself, in a very tongue-in-cheek manner:
This one may be called the dominant, the other one the recessive anlage [ed: Correns’s word for ‘allele’ (part of a gene)]. Mendel named them in this way, and, by a strange coincidence, de Vries now does likewise.
Of the three men, de Vries certainly had the most to lose, as his reputation was the most established. We may never know whether or not de Vries or either of the others formulated their own versions of Mendel’s laws, but whatever the case may be, after 35 years of languishing, Mendel’s work was suddenly on everyone’s mind. And the time was ripe for someone to come along and connect Mendel and Weismann and usher in a new age of genetics research.
The magic number
The connection was made by an American graduate student named Walter Sutton, who was performing his doctoral research at Columbia University under the famous zoologist Edmund Beecher Wilson. Sutton had recently read a paper by a German professor named Theodor Boveri describing several experiments with sea urchin embryos. In the course of his work, Boveri developed the intuition that sea urchin embryos required a specific number of chromosomes to develop normally. Too many or too few and the embryo would die before it developed significantly. Boveri had observed that roughly equal numbers of chromosomes come together at fertilization to give a viable embryo. This was easy enough to see if one fixed and stained the fertilized eggs. However, in doing so, the eggs were killed. Boveri wanted to show that this fact still held in living eggs. So he came up with a remarkably elegant solution to observe what happened to living sea urchin embryos that had the wrong number of chromosomes: he used sea urchin eggs which had been fertilized with two sperm simultaneously. These so-called dispermic eggs did not need to be fixed and stained: it was obvious that they did not have the correct number of chromosomes. Boveri’s hypothesis was that the embryos would not develop normally. And they didn’t: the mis-fertilized eggs died in droves. The experiment had shown that chromosomes must come together in a specific number during fertilization.
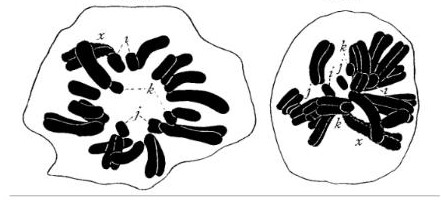
Sutton was working on embryo development in grasshoppers at the time. This provided him with a distinct, if fortuitous, advantage: the chromosomes in the grasshopper were easily visible and varied in size significantly enough for Sutton to be able to distinguish between individual chromosomes (see above pictures). With Boveri’s results in hand, Sutton confirmed that in grasshoppers as well as sea urchins, it was necessary for one set of chromosomes each from sperm and egg to come together to give the correct number and distribution for proper development of embryos. He was aware that this behavior of chromosomes was eerily similar to the behavior predicted by genes in Mendel’s laws of inheritance. At the end of his 1902 paper, he hinted at the importance of his result:
I may finally call attention to the probability that the association of paternal and maternal chromosomes in pairs and their subsequent separation during the reducing division [ed: ‘reducing division’ is Sutton’s term for ‘meiosis’] as indicated above may constitute the physical basis of the Mendelian law of heredity. To this subject I hope soon to return in another place.
Sutton would release a more extensive paper the following year where he laid out the most important result in genetics up to that point: Chromosomes are the carriers of genes. This contribution to the field was important enough that we still refer to the chromosome theory of inheritance as the Boveri-Sutton theory.
Bright eyes
The Boveri-Sutton theory was as controversial as it was important. Immediately, defenders and detractors sprung up around the theory and its originators. One of the main opponents of the theory was an American named Thomas Hunt Morgan. Morgan’s main bone of contention was the observation of linked genes. A little background is in order. In 1905, a group of British scientists (William Bateson, Edith Rebecca Saunders, and Reginald Punnett) were examining pea plants in the same way that Mendel had done back in the 1860’s. They were looking specifically at flower color and pollen grain shape. In this species of pea, the flowers were either red or purple and the pollen grains were either long or short. The three noticed that plants with purple flowers had long pollen grains far more often than they had short pollen grains. But this shouldn’t have been the case. One of Mendel’s laws states that genes are inherited independently of one another. But here, the ratios of offspring were not at all what Mendel’s laws predicted. It looked like the “pollen grain shape” gene and the “flower color” gene were being inherited together. These two genes were referred to as linked genes.
There was a nascent theory going around that genetic linkage could be explained if the two genes were physically on the same chromosome–that way, they’d both be sorted together during meiosis and both be picked up during fertilization. This theory preserved the chromosome theory of heredity, but there were problems with it. For instance, in the case of Bateson, Saunders, and Punnett’s experiments, the purple flowers usually had long pollen grains, but the group still observed some plants with red flowers and long pollen grains, or purple flowers and short pollen grains. If these observations were correct, the genes must not be completely linked. But then if they’re not completely linked, then either the genes aren’t on the same chromosome, or Mendel’s laws are wrong. In fact, there’s another explanation that we’ll get to in a minute that allows us to keep the chromosome theory and Mendel’s laws, but the observation of not-completely-linked genes dealt a serious blow to the chromosome theory of inheritance, and to Mendel.
Morgan thought the idea of linked genes was just a case of fiddling with the theory until it fit the data. It was all too speculative, but being a scientist, Morgan set out to test inheritance theories for himself. So he set up a lab at Columbia University (just down the hall from where Wilson and Sutton were working), where he and his students began to study a new model organism: Drosophila Melanogaster, or as we know it better, the fruit fly.
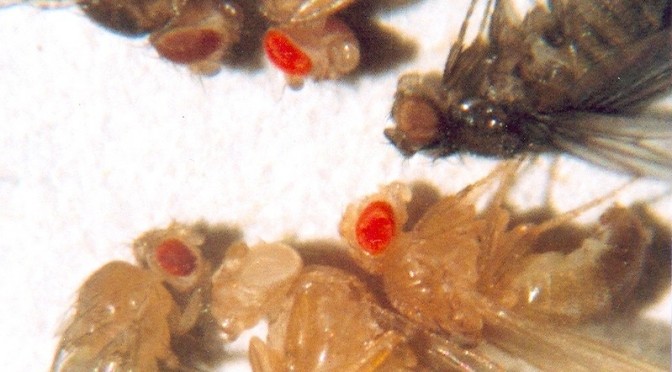
Fruit flies proved to be the perfect organism to carry out genetic experiments: they reproduced and carried out their life cycles quickly (average lifespan was around a month), and they were cheap, small, and easily obtainable–important considerations when lab space and funding was scarce. Morgan’s group began blasting generation after generation of hapless fruit flies with “wide ranges of temperature, salts, sugars, acids, alkalis, and radium and X rays,” hoping to induce some sort of mutation. Many mutations were observed, but they were all discarded in the belief that the mutations were naturally occurring, and not artificially induced. Finally, in 1910, Morgan noticed something odd: a male fruit fly with white eyes. Typically, fruit flies have bright red-orange eyes–indeed, since the flies are so small, their bright eyes are usually the only things you can make out without the aid of a microscope. So this white-eyed male stood out as anomalous.

Morgan began breeding the white-eyed male with females, all of whom had the common red-orange eye color. Their offspring all had red eyes, but Morgan pressed on: if Mendel’s laws are true, then cross-breeding this generation of flies should produce white-eyed flies in a 3:1 ratio, as we explained last time. This was, in fact, what he saw. But there was a catch: all of the white-eyed fruit flies were male. At first, it was believed that the white-eyed gene might kill female embryos, but when Morgan crossed white-eyed males with the female offspring of whites and reds, he got white-eyed females and males. Eventually, he turned his attention to the theory he had railed against. Wilson and Sutton, along with their co-worker Nettie Stevens, had produced cell observations that very strongly suggested that sex determination–whether an animal is male or female–was governed by a pair of chromosomes which would eventually be called X and Y. If an organism had two X’s, it would be female, and if it had an X and a Y, it would be male. Morgan began to examine what would happen if the white-eyed gene was linked to the sex chromosomes, and if Mendel’s laws were correct after all. Assuming that the white-eyed gene is recessive, and that the white-eyed gene only occurred on the X chromosome, and not on the Y chromosome, Morgan worked out what would happen in each of his experiments:
1) If a white-eyed male mates with a normal red-eyed female:
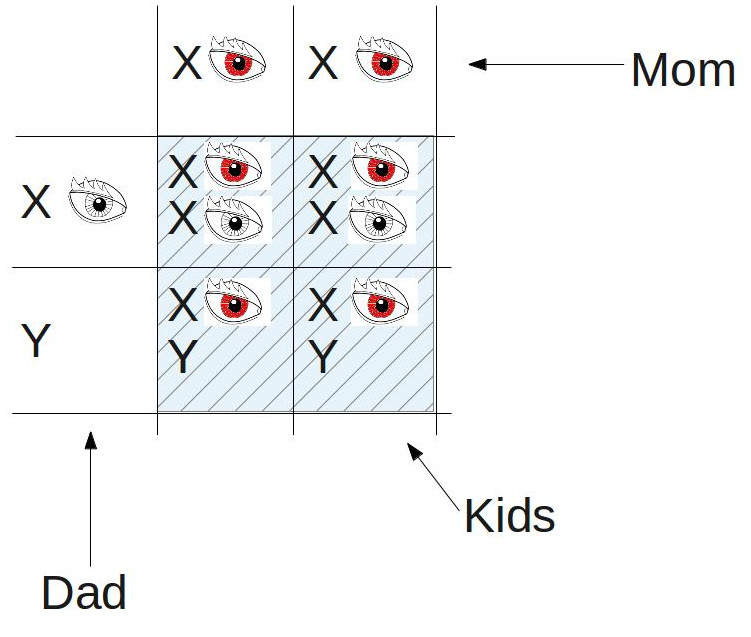 Since red eyes are dominant over white eyes, all the kids will have red eyes. But notice something: remember that the gene for eye color is only on the X chromosome. Since females have two X’s, they can be carriers for the white-eyed gene without displaying white eyes. But since males only have one X, they display whatever eye color gene they receive from their mothers. You can see above that both the male kids (bottom row) have the gene for red eyes that they inherited from their mother. So we move on to the second experiment:
Since red eyes are dominant over white eyes, all the kids will have red eyes. But notice something: remember that the gene for eye color is only on the X chromosome. Since females have two X’s, they can be carriers for the white-eyed gene without displaying white eyes. But since males only have one X, they display whatever eye color gene they receive from their mothers. You can see above that both the male kids (bottom row) have the gene for red eyes that they inherited from their mother. So we move on to the second experiment:
2) If a red-eyed male mates with one of the female kids from the square above:
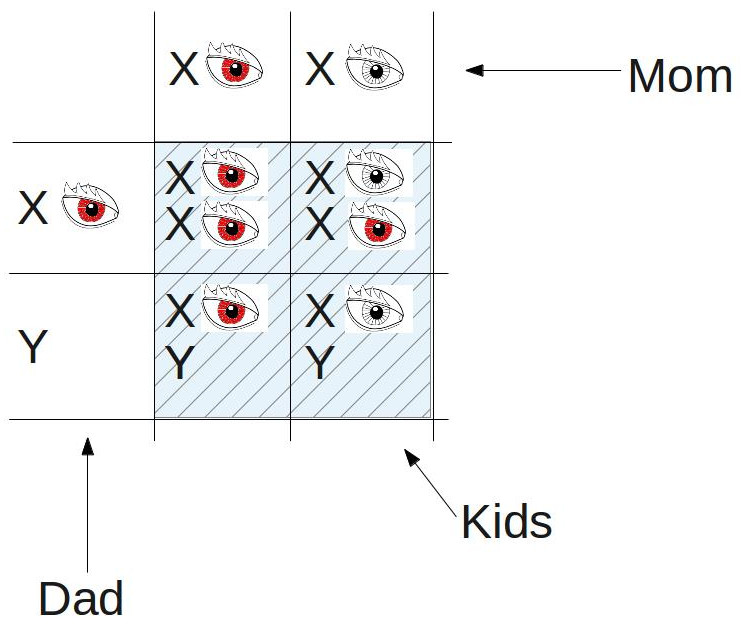 The ratio of red-eyed kids to white-eyed kids is 3:1, just as Mendel would have predicted, but, as we see above, all of the white-eyed kids are male. The evidence is growing stronger: it looks like the gene for eye color very well may reside on the X chromosome. And then came Morgan’s master stroke: he mated a white-eyed male with a female offspring from his first experiment above…
The ratio of red-eyed kids to white-eyed kids is 3:1, just as Mendel would have predicted, but, as we see above, all of the white-eyed kids are male. The evidence is growing stronger: it looks like the gene for eye color very well may reside on the X chromosome. And then came Morgan’s master stroke: he mated a white-eyed male with a female offspring from his first experiment above…
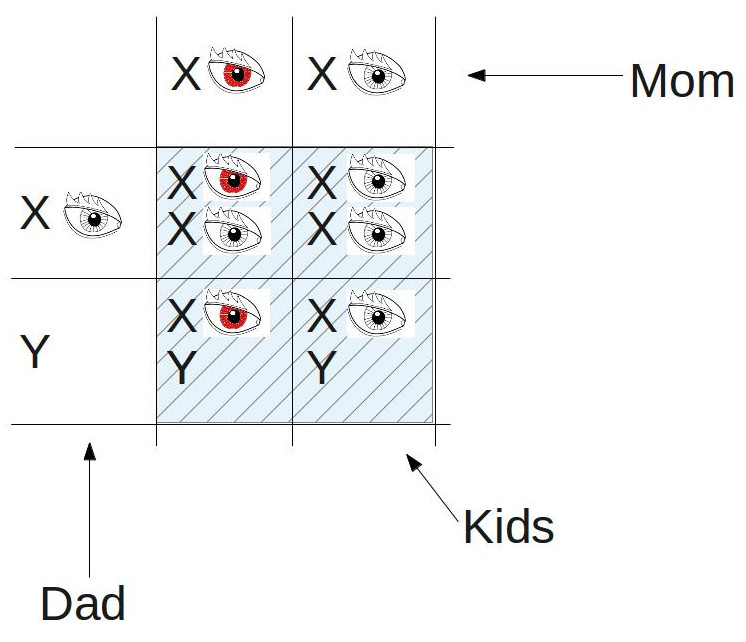 …showing that it was possible, albeit unlikely, for a female to be born with white eyes. This was the experiment that convinced Morgan of the veracity of the chromosome theory of inheritance, and of Mendel’s place within the theory.
…showing that it was possible, albeit unlikely, for a female to be born with white eyes. This was the experiment that convinced Morgan of the veracity of the chromosome theory of inheritance, and of Mendel’s place within the theory.
Crossing over
Of course, there was a snag. Eye color was the first and most famous sex-linked trait that Morgan observed, but there was another important one. He observed a mutation that caused the flies’ wings to be underdeveloped, and noticed that it obeyed the same patterns of inheritance that the white eyes did: it was linked to the sex of the fruit fly. This meant that both the eye color gene and the wing size gene both resided on the X chromosome. Since they were both on the same chromosome, they were linked genes, just like the flower color and pollen shape in the pea plants of Bateson, Saunders, and Punnett. But Morgan noticed, as we mentioned earlier, that the genes were not-completely-linked. That is to say, they were often, but not always, inherited together. How could the chromosome theory be saved, now that Morgan was a recent convert?
In 1909, Belgian named Frans Alfons Janssens had observed an odd phenomenon in cell experiments. Right at the very beginning of meiosis, pairs of chromosomes appeared to cross and exchange pieces. Here’s a schematic that Morgan put together to describe what was going on:
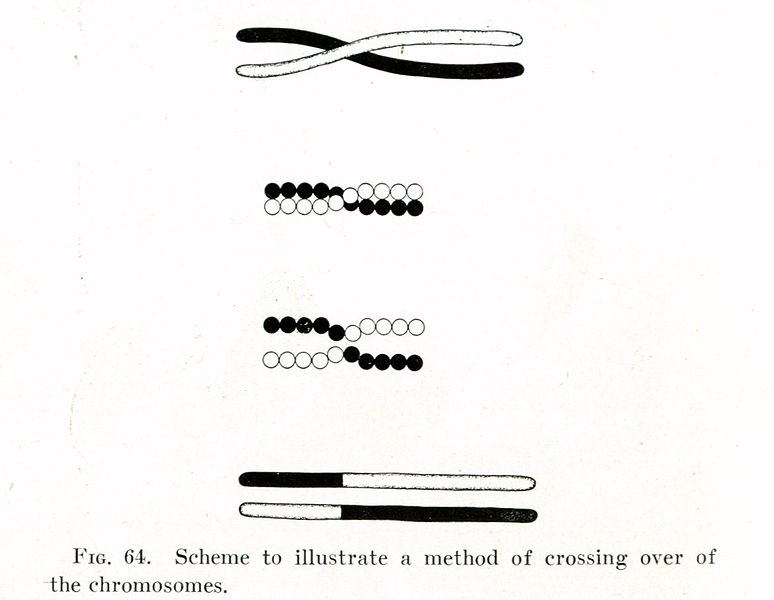
In this illustration, one chromosome in a pair (white) crosses over its partner (black). During the process, part of the white chromosome is transferred onto the black chromosome. Morgan realized that this was the answer to his not-completely-linked genes conundrum. Since linked genes were believed to reside on the same chromosome, they would have to be inherited together. But if this crossing over process exchanged pieces of chromosomes, then this provided a way for genes to become “unlinked.” So with the addition of crossing over, the chromosome theory of inheritance and Mendel’s laws were finally vindicated.
There’s another trick that geneticists can play by looking at linked genes and crossing over. Let’s look at Morgan’s diagrams again, but this time, we’ll label four hypothetical genes on the chromosomes:
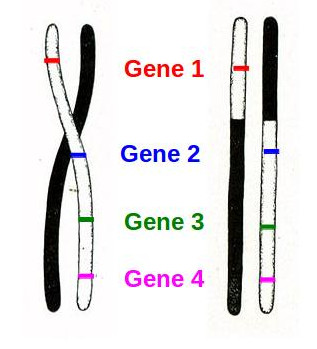 It’s pretty clear that if crossing over occurs at a random position somewhere roughly in the middle of the chromosomes, that gene 1 and gene 4 are more likely to end up on different chromosomes than, say, gene 3 and gene 4. This means that the closer together on the actual chromosome two genes are, the more they will appear to be linked in analyzing offspring. In fact, if genes are very strongly linked (like flower color and pollen grain shape in Bateson’s pea plants), this is a good indication that they are very close together on the chromosome. For less closely linked genes (like the white eyes and underdeveloped wings in Morgan’s fruit flies), this indicates that they are farther apart on the chromosome. Geneticists can use the statistics of how often these traits appear in large numbers of offspring of breeding experiments to map out roughly where genes occur on chromosomes. In fact, Alfred Sturtevant, a graduate student of Morgan, did just that in 1913, working out the placement of six separate genes on the X chromosome of fruit flies.
It’s pretty clear that if crossing over occurs at a random position somewhere roughly in the middle of the chromosomes, that gene 1 and gene 4 are more likely to end up on different chromosomes than, say, gene 3 and gene 4. This means that the closer together on the actual chromosome two genes are, the more they will appear to be linked in analyzing offspring. In fact, if genes are very strongly linked (like flower color and pollen grain shape in Bateson’s pea plants), this is a good indication that they are very close together on the chromosome. For less closely linked genes (like the white eyes and underdeveloped wings in Morgan’s fruit flies), this indicates that they are farther apart on the chromosome. Geneticists can use the statistics of how often these traits appear in large numbers of offspring of breeding experiments to map out roughly where genes occur on chromosomes. In fact, Alfred Sturtevant, a graduate student of Morgan, did just that in 1913, working out the placement of six separate genes on the X chromosome of fruit flies.
Conclusion
We’ve come a long way in this post. We started off with Mendel in relative obscurity and Weismann with scant support of his theory. In the space of only 13 years, we rediscovered Mendel, pieced together that chromosomes carry genes that obey Mendel’s laws, discovered that gender is mediated by chromosomes, explained linked genes and chromosomal crossing over, and began mapping out genes on chromosomes. We’ve only begun to scratch the surface. As frantic as the period from 1900-1913 was for genetics, it was just as frantic for biochemistry starting in the mid-1920’s. But we’ll have to save that story for the next chapter. Stay tuned.
References
There are many, many papers debating the specific details of the rediscovery of Mendel’s laws: who should be given credit, how much of their work was their own and how much was Mendel’s, etc. Here are a few:
- Kottler, M.J. “Hugo de Vries and the rediscovery of Mendel’s laws.” Annals of Science. 1979, 36, 517.
- Simunek, M.; Hoßfeld, U.; and O. Breidbach. “‘Further Development’ of Mendel’s legacy? Erich von Tschermak-Seysenegg in the context of Mendelian–biometry controversy, 1901–1906.” Theory in Biosciences 2012, 131, 243.
- Harwood, J. “The rediscovery of Mendelism in agricultural context: Erich von Tschermak as plant-breeder.” Comptes Rendus de l’Académie des Sciences – Series III – Sciences de la Vie. 2000, 323, 1061.
- Corcos, A.; Monaghan, F. “Role of de Vries in the recovery of Mendel’s work. I. Was de Vries really an independent discoverer of Mendel?” Journal of Heredity. 1985, 76, 187.
Here are links to the English translations of the papers by de Vries, Correns, and Tschermak:
- De Vries: http://www.esp.org/foundations/genetics/classical/holdings/v/hdv-00.pdf
- Correns: http://www.esp.org/foundations/genetics/classical/holdings/c/cc-00.pdf
- Tschermak: http://www.esp.org/foundations/genetics/classical/holdings/t/et-00.pdf
Sutton’s papers are here:
- http://www.esp.org/foundations/genetics/classical/wss-02.pdf
- http://www.esp.org/foundations/genetics/classical/holdings/s/wss-03.pdf
Incidentally, this is the first time I’m realizing how many of these papers are from esp.org (Electronic Scholarly Publishing). It’s a really fantastic resource for classic papers in genetics.
Boveri’s sea urchin experiments are described in detail here:
An article on genetic linkage from Nature Education:
- Lobo, I. & Shaw, K. “Discovery and types of genetic linkage.” Nature Education. 2008.
A memoir on Morgan by his student, Alfred Sturtevant, can be found here:
The Punnett squares in the fruit fly section are adapted from this article:
- Miko, I. “Thomas Hunt Morgan and sex linkage.” Nature Education. 2008.